Preeclampsia (PE) is a multisystem disorder, that affects approximately 5 % of
pregnancies worldwide. PE is one of the leading causes of maternal, fetal and neonatal
morbidity or mortality. The onset of PE occurs about 20. gestational week (exceptionally
before 20 weeks; Tanaka et al., 2015) and it is characterized by many heterogenous
symptoms. The most detectable symptoms are hypertension and proteinuria, which also
serves as the main diagnostic markers. In addition, several other symptoms could be linked to
PE such as abnormalities of placenta, kidney, and liver, neurological and hematological
complications which could lead to fetal damage. In general, the treatment of PE is to manage
the symptoms however the disorder persists. Since PE is a disorder of the placenta, the only
direct treatment is the delivery. More than half of women are forced to undergo the delivery
in the form of a section (Roberts and Lain, 2002; Zhang et al, 2003; Wagner, 2004; Liu et al.,
2009).
Diagnostic criteria
The major criteria for diagnosing the disease are de novo hypertension, which appears
after 20 weeks gestation, in combination with proteinuria. Preeclamptic hypertension is
defined as systolic blood pressure ≥ 140 mm Hg and/or diastolic blood pressure ≥ 90 mm Hg,
measured twice, at intervals of 4 – 6 hours. Proteinuria (elevated levels of proteins in urine) is
determined by the protein content in urine ≥ 300 mg/day (Mol et al., 2016; Townsend et al.,
2016).
There is a group of abnormalities, known together as the hypertensive disorders of
pregnancy, which includes preeclampsia and gestational hypertension. It is possible, that
hypertension is elevated for the first time after 20 weeks gestation, suggesting a potential
onset of PE, but the patient does not suffer from proteinuria. This condition is classified as
having gestational hypertension. One of four women, suffering from gestational
hypertension, subsequently develop proteinuria, leading to the onset of PE (Saudan et al.,
1998; Report of the National High Blood Pressure Education Program Working Group on
High Blood Pressure in Pregnancy, 2000; Wagner, 2004). The similarity of symptoms
detected in the hypertensive disorders of are the reason of the unclear and inaccurate
diagnostics of PE.
Dong et al. monitored PE in women of varying severity of proteinuria. Based on the
obtained data, authors suggest that the level of proteins in urine may be associated with the
development of severity of PE. In addition, they refer, that the amount of proteinuria is
positively correlated with time of onset in PE, but time between the onset of PE and delivery
is not associated with the severity of proteinuria (Dong et al., 2017). Despite the mentioned
diagnostic features, up to 10 % of patients with PE have no proteinuria at the time of clinical
presentation (Sibai, 1990; Thornton et al., 2010; Dong et al., 2017). These knowledges
complicate the detection of disorder onset causing difficulties in diagnostics of PE.
Trophoblast and placenta
The structure and functionality of the placenta is related to the health of mother and
developing fetus (Guttmacher et al., 2014). The process of placentation is the key point in PE,
because the placenta formation provides the functional connection between mother and fetus.
The result of placentation is the implantation of blastocyst in uterus. Invasion of the
trophoblast into the maternal endometrium is necessary for placentation. The blastocyst
contains a layer of cytotrophoblast cells on its surface, the cells that fuse and form a
multinucleated syncytiotrophoblast (fig. 1). Only the multinucleated unit can invade into the
maternal endometrium, what is indispensable for the proper placentation process (Popek,
1999; Ji et al., 2013).
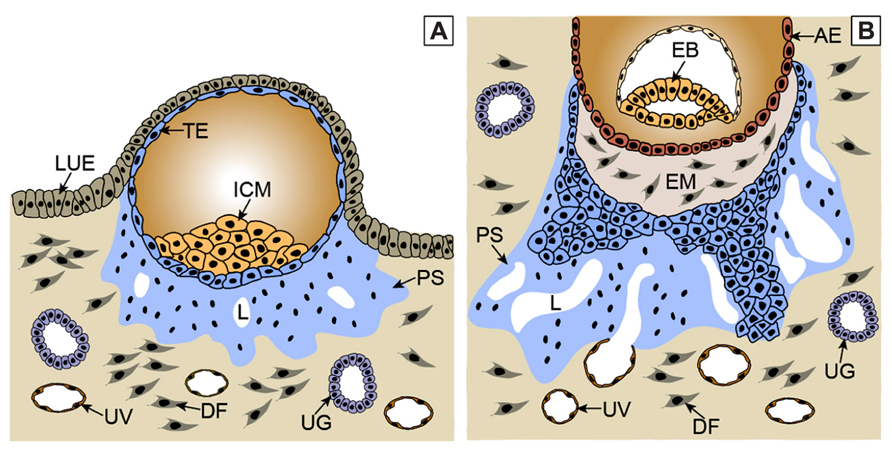
Figure 1: Trophoblast invasion processA: After the implantation, the trophoectoderm stem cells give rise to primitive multinucleated syncytium by
fusing. In the region of lacunae, the ancestor of intervillar space is formed. Some of the lacunae erode
uterine vessels.
B: In the next stage, the cells of proliferative cytotrophoblast are flowing from the trophoectoderm, break
through the primitive syncytium and form primary villi.
AE – amniotic epithelium; DF – decidual fibroblast; EB – embryoblast; EM – extraembryonic mesoderm;
ICM – inner cell mass; LUE – luminal uterine epithelium; L – lacunae; PS – primitive syncytium; TE –
trophoectoderm; UG – uterine gland; UV – uterine vessel (Knöfler and Pollheimer, 2013).
Syncytin-1 in placentation
Syncytin-1 protein is encoded by ERVW-1 gene belonging to the human endogenous
retroviral family – HERV-W. The highest level of ERVW-1 expression was detected in
placenta. The product of this gene mediates the cytotrophoblast cell fusion, from which
differentiates multinucleated syncytiotrophoblast (fig. 2). Without the fusogenic activity of
syncytin-1, normal placentation could not occur (Blond et al., 1999; Blond et al., 2000; Mi et
al., 2000; Voisset et al, 2000; Huang et al., 2014).
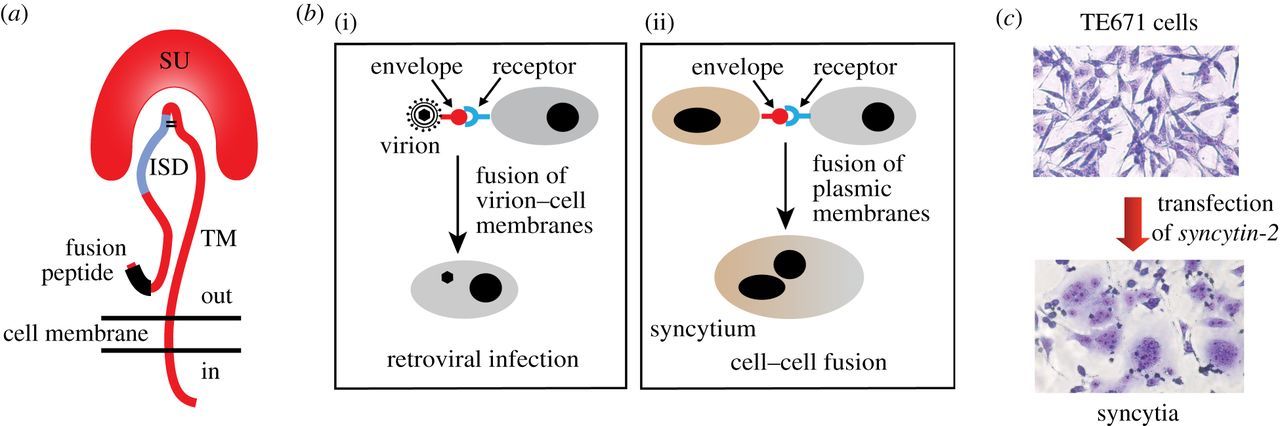
Figure 2: The retroviral envelope glycoprotein. (a) Structure of the retroviral envelope protein with the SU
and TM subunits, the fusion peptide and the immunosuppressive domain (ISD). (b) Consequences of the
interaction between a retroviral envelope protein and its receptor: virion–cell membrane fusion and virus
entry into the target cell (i) or cell–cell membrane fusion and formation of a syncytium (ii). (c) Cell–cell
fusion and formation of multinucleated syncytia induced by transfecting human TE671 cells with a syncytin-
2 expression vector (May–Grünwald–Giemsa staining) (Lavialle et al., 2013)
Molecular structure of ERVW-1 gene
ERVW-1 gene is located on q21.2 of chromosome 7 in the genome. It consists of
three, originally retroviral, parts of which gag and pol regions are found in the human
genome as pseudogenes, while the env (coding envelope protein) region has been
functionally preserved. Only the env region has retained complete retroviral ORF (Open
Reading Frame), whose product is fusogenic protein syncytin-1, required for the cell fusion
in syncytiotrophoblast forming (fig. 3). Env sequence is flanked by two LTR (Long Terminal
Repeat) regions. Downstream from 5ʼLTR is laying an intron with the size of 2 kb (Blond et
al, 1999; Blond et al., 2000; Voisset et al., 2000).
The structure of the syncytin-1 protein
The functional syncytin-1 consist of surface-associated unit (SU) and transmembrane
unit (TM), which are linked to each other by disulfide bonds. SU plays a role in interaction
with receptors. The role of the TM is to cross the membrane and anchor the peptide in the cell
membrane. Moreover, TM contains the potential immunosuppressive domain, which has a
competence to supress the immune response of mother towards fetus. Fully functional
syncytin-1 is organised as a homotrimeric structure. Without this spatial arrangement, it loses
the fusogenic activity (Fass and Kim, 1995; Cheynet et al., 2005).
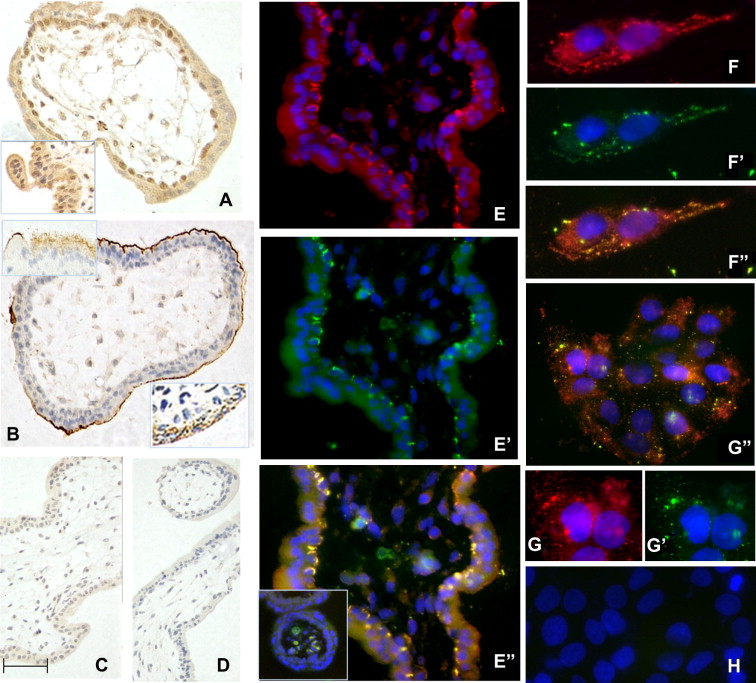
Figure 3: Immunolocalisation of syncytin 1 in first trimester placenta (A–E) and BeWo cells (F–H). (A)
Surface (SU) subunit; (B) transmembrane (TM) subunit; (C) IgG control for SU; (D) IgG control for TM.
DAB staining, with haematoxylin counterstain. (E) Dual immunofluorescence localisation of syncytin 1
subunits in placental tissue probed with antibodies targeting SU (red) or TM (green). (F, G) BeWo cells
probed with antibodies targeting SU (red) or TM (green). Co-localisation of the two subunits is seen in
yellow (F″, G″). H shows a negative control. DAPI nuclear counterstain (blue). The scale bar shown in C
corresponds to: A, B:18 μm; C, D:25 μm; E: 10 μm; F, G″, H: 15 μm; G,G′: 8 μm.
Syncytin-1 and preeclampsia
According to Holder et al., the level of syncytin-1 is elevated in placenta of PE
women, compared to physiological placentas (Holder et al., 2012). This founding contrasts
with other studies, they refer decreased expression of syncytin-1 at the mRNA but also
protein level (Lee et al., 2001; Knerr et al., 2002; Vargas et al., 2011). In addition, many
studies included inadequate number of samples considering the frequency of incidence of PE.
Despite the knowledge obtained regarding the association between syncytin-1 and PE, the
overall conclusions are still unclear. Consequently, there is a need for a closer study of the
pathophysiology of PE in terms of correlation of ERVW-1 gene expression with the disease,
and the results need to be demonstrated in a sufficiently large sample of patients.
References
Blond, J. L., Besème, F., Duret, L., Bouton, O., Bedin, F., Perron, H., Mandrand., B., Mallet, F.
(1999). Molecular characterization and placental expression of HERV-W, a new human endogenous
retrovirus family. J Virol.; 73(2): 1175–1185.
Blond, J. L., Lavillette, D., Cheynet, V., Bouton, O., Oriol, G., Chapel-Fernandes, S., Mandrand, B.,
Mallet, F., Cosset, F. L. (2000). An envelope glycoprotein of the human endogenous retrovirus
HERV-W is expressed in the human placenta and fuses cells expressing the type D mammalian
retrovirus receptor. J Virol.; 74(7): 3321–3329.
Buchrieser, J., Degrelle, S. A., Couderc, T., Nevers, Q., Disson, O., Manet, C., Donahue, D. A.,
Porrot, F., Hillion, K. H., Perthame, E., Arroyo, M. V., Souquere, S., Ruigrok, K., Dupressoir, A.,
Heidmann, T., Montagutelli, X., Fournier, T., Lecuit, M., & Schwartz, O. (2019). IFITM proteins
inhibit placental syncytiotrophoblast formation and promote fetal demise. Science; 365(6449):
176–180.
Dong, X., Gou, W., Li, C., Wu, M., Han, Z., Li, X., Chen, Q. (2017). Proteinuria in preeclampsia: Not
essential to diagnosis but related to disease severity and fetal outcomes. Pregnancy Hypertens; 8:60-
64.
Fass, D., Kim, P. S. (1995). Dissection of a retrovirus envelope protein reveals structural similarity
to influenza hemagglutinin. Curr. Biol.; 5: 1377–1383.
Frendo, J. L., Olivier, D., Cheynet, V., Blond, J. L., Bouton, O., Vidaud, M., Rabreau, M., Evain-
Brion, D., Mallet, F. (2003). Direct involvement of HERV-W Env glycoprotein in human trophoblast
cell fusion and differentiation. Mol. Cell. Biol; 23(10): 3566–3574.
Guttmacher, A. E., Maddox, Y. T., Spong, C. Y. (2014). The Human Placenta Project: placental
structure, development, and function in real time. Placenta; 35(5): 303–4.
Holder, B. S., Tower, C. L., Abrahams, V. M., Aplin J. D. (2012). Syncytin 1 in the human
placenta. Placenta; 33(6): 460–466.
Huang, Q., Chen, H., Li, J., Oliver, M., Ma, X., Byck, D., Gao, Y., Jiang, S. W. (2014). Epigenetic
and non-epigenetic regulation of syncytin-1 expression in human placenta and cancer tissues. Cell
Signal.; 26(3): 648–56.
Cheynet, V., Ruggieri, A., Oriol, G., Blond, J. L., Boson, B., Vachot, L., Verrier, B., Cosset, F. L,
Mallet, F. (2005). Synthesis, assembly, and processing of the Env ERVWE1/syncytin human
endogenous retroviral envelope. J Virol.; 79(9): 5585–93.
Chiang, M. H., Liang, F. Y., Chen, C. P., Chang, C. W., Cheong, M. L., Wang, L. J., Liang, C. Y.,
Lin, F. Y., Chou, C. C., & Chen, H. (2009). Mechanism of hypoxia-induced GCM1 degradation:
implications for the pathogenesis of preeclampsia. J Biol Chem; 284(26): 17411–17419.
Ji, L., Brkić, J., Liu, M., Fu, G., Peng, C., Wang, Y. L. (2013). Placental trophoblast cell
differentiation: physiological regulation and pathological relevance to preeclampsia. Mol Aspects
Med.; 34(5): 981–1023.
Knerr, I., Beinder, E., Rascher, W. (2002). Syncytin, a novel human endogenous retroviral gene in
human placenta: evidence for its dysregulation in preeclampsia and HELLP syndrome. Am J Obstet
Gynecol;186(2): 210–3.
Knöfler, M. a Pollheimer, J. (2013). Human placental trophoblast invasion and differentiation: a
particular focus on Wnt signaling. Front Genet.; 4: 190.
Lee, X., Keith, Jr. J. C., Stumm, N., Moutsatsos, I., McCoy, J. M., Crum, C. P., Genest, D., Chin,
D., Ehrenfels, C., Pijnenborg, R., van Assche, F. A., Mi, S. (2001). Downregulation of placental
syncytin expression and abnormal protein localization in pre-eclampsia. Placenta; 22(10): 808–12.
Liu, A., Wen, S. W., Bottomley, J., Walker, M. C., Smith, G. (2009). Utilization of Health Care
Services of Pregnant Women Complicated by Preeclampsia in Ontario. Hypertens Pregnancy;
28(1): 76–84.
Mi, S., Lee, X., Li, X., Veldman, G. M., Finnerty, H., Racie, L., LaVallie, E., Tang, X. Y., Edouard,
P., Howes, S., Keith, J. C. J., McCoy J. M. (2000). Syncytin is a captive retroviral envelope protein
involved in human placental morphogenesis. Nature; 403: 785–788.
Mol, B. W. J., Roberts, C. T., Thangaratinam, S., Magee, L. A., de Groot, C. J. M., Hofmeyr, G. J.
(2016). Pre-eclampsia. Lancet; 387(10022): 999–1011.
Popek, E. J., (1999). Normal anatomy and history of the placenta. In: Pathology of the Placenta.
Lewis, S. H., Perrin, E. Harcourt Brace & Company; 49–88.
Report of the National High Blood Pressure Education Program Working Group on High Blood
Pressure in Pregnancy (2000). Am J Obstet Gynecol; 183(1).
Roberts, J. M, Lain, K. Y. (2002). Recent insights into the pathogenesis of pre-eclampsia. Placenta;
23(5): 359–372.
Saudan, P., Brown, M. A., Buddle, M. L., Jones, M. (1998). Does gestational hypertension become
pre-eclampsia? Br J Obstet Gynaecol.; 105(11): 1177-84.
Sibai, B. M. (1990). Eclampsia. VI. Maternal-perinatal outcome in 254 consecutive
cases. Am J Obstet Gynecol; 163(3): 1049–1054.
Soygur, B., & Sati, L. (2016). The role of syncytins in human reproduction and reproductive organ
cancers, Reproduction; 152(5): R167-78.
Tanaka, M., Tsujimoto, Y., Goto, K., Kumahara, K., Onishi, S., Iwanari, S., Fumihara, D., Miki,
S., Ikeda, M., Sato, K., Sato, H., Hirose, M., Takeoka, H. (2015). Preeclampsia before 20 weeks of
gestation: a case report and review of the literature. CEN Case Rep.; 4(1): 55–60.
Thornton, C. E., Makris, A., Ogle, R. F., Tooher, J. M., Hennessy, A. (2010). Role of proteinuria in
defining pre‐eclampsia: Clinical outcomes for women and babies. Clin. Exp. Pharmacol. Physiol.;
37(4): 466-470.
Tolosa, J. M., Schjenken, J. E., Clifton, V. L., Vargas, A., Barbeau, B., Lowry, P., Maiti, K., & Smith,
R. (2012). The endogenous retroviral envelope protein syncytin-1 inhibits LPS/PHA-stimulated
cytokine responses in human blood and is sorted into placental exosomes. Placenta; 33(11): 933–941.
Townsend, R., O'Brien, P., Khalil, A. (2016). Current best practice in the management of
hypertensive disorders in pregnancy. Integr Blood Press Control; 9: 79–94.
Vargas, A., Toufaily, C., Lebellego, F., Rassart, E., Lafond, J., Barbeau, B. (2011). Reduced
expression of both syncytin 1 and syncytin 2 correlates with severity of preeclampsia. Reprod Sci;
18(11): 1085–91.
Voisset, C., Bouton, O., Bedin, F., Duret, L., Mandrand, B., Mallet, F., Paranhos-Baccalà, G.
(2000). Chromosomal Distribution and Coding Capacity of the Human Endogenous Retrovirus
HERV-W Family. AIDS Res Hum Retroviruses, 16(8): 731–740.
Wagner, L. K. (2004). Diagnosis and Management of Preeclampsia. Am Fam Physician; 70(12):
2317–2324.
Wang, X., Liu, Z., Wang, P., Li, S., Zeng, J., Tu, X., Yan, Q., Xiao, Z., Pan, M., Zhu, F. (2018).
Syncytin-1, an endogenous retroviral protein, triggers the activation of CRP via TLR3 signal cascade
in glial cells. Brain Behav Immun; 67: 324–34.
Zhang, J., Meikle, S., Trumble, A. (2003). Severe maternal morbidity associated with hypertensive
disorders in pregnancy in the United States. Hypertens Pregnancy; 22(2): 203–212.
Zhu, H., Peng, B., Klausen, C., Yi, Y., Li, Y., Xiong, S., von Dadelszen, P., & Leung, P. (2020).
NPFF increases fusogenic proteins syncytin 1 and syncytin 2 via GCM1 in first trimester primary
human cytotrophoblast cells. FASEB J; 34(7): 9419–9432.
Project supported by the Ministry of Health of the Slovak Republic (MZSR 2018/40-LFUK-14)
Supported priority areas chosen from the approved list for a year 2018: Innovative diagnostic and
therapeutic procedures and products personalized / precision medicine - 1. Early, fast and valid
diagnosis - Products for the identification of novel biomarkers of disease